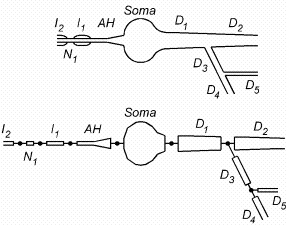
|
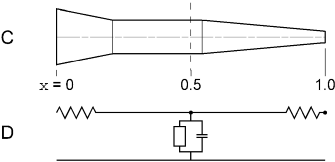
 Distance along the length of a section is discussed in terms of the normalized
position parameter x. That is, one end of the section
corresponds to x = 0 and the other end to x = 1. In Fig.
3.3C these locations are depicted as being on the left and right hand ends of
the section. The locations of the nodes and the boundaries between segments
are conveniently specified in terms of this normalized position parameter. In
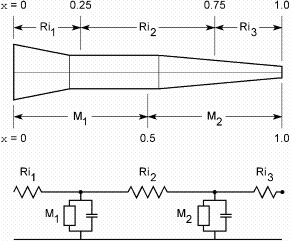
general, a section has nseg segments that are demarcated by evenly
spaced boundaries at intervals of 1 / nseg. The nodes at the centers
of these segments are located at x = (2 i - 1) / 2
nseg where i is an integer in the range [1, nseg]. As
we shall see later, x is also used in specifying model parameters or
retrieving state variables that are a function of position along a section (see
4.4 Range variables).
Distance along the length of a section is discussed in terms of the normalized
position parameter x. That is, one end of the section
corresponds to x = 0 and the other end to x = 1. In Fig.
3.3C these locations are depicted as being on the left and right hand ends of
the section. The locations of the nodes and the boundaries between segments
are conveniently specified in terms of this normalized position parameter. In
general, a section has nseg segments that are demarcated by evenly
spaced boundaries at intervals of 1 / nseg. The nodes at the centers
of these segments are located at x = (2 i - 1) / 2
nseg where i is an integer in the range [1, nseg]. As
we shall see later, x is also used in specifying model parameters or
retrieving state variables that are a function of position along a section (see
4.4 Range variables).
The special importance of x and nseg lies in the fact that
they free the user from having to keep track of the correspondence between
segment number and position on the nerve. In early versions of NEURON, all
nerve properties were stored in vector variables where the vector index was the
segment number. Changing the number of segments was an error prone and
laborious process that demanded a remapping of the relationship between the
user's mental image of the biologically important features of the model, on the
one hand, and the implementation of this model in a digital computer, on the
other. The use of x and nseg insulates the user from the
most inconvenient aspects of such low-level details.
When nseg = 1 the entire section is lumped into a single compartment.
This compartment has only one node, which is located midway along its length,
i.e. at x = 0.5 (Fig. 3.3C and D). The integral of the surface area
over the entire length of the section (0 <= x <= 1) is used to
calculate the membrane properties associated with this node. The values of the
axial resistors are determined by integrating the cytoplasmic resistivity along
the paths from the ends of the section to its midpoint (dashed line in Fig.
3.3C). The left and right hand axial resistances of Fig. 3.3D are evaluated
over the x intervals [0, 0.5] and [0.5, 1], respectively.
|