|
|
|
|
|
|
|
|
(1) |
| which describes the relationship between current and voltage in a one-dimensional cable. The branched architecture typical of most neurons is incorporated by combining equations of this form with appropriate boundary conditions. |
|
|
|
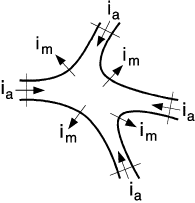
| Figure 3.1. The net current entering a region must equal zero. |
|
|
|
|
|
(2) |
|
|
|
|
|
|
If the compartments have the same length
|
|
|
|
|
where we have replaced the total ionic current
|
|
|
|
Address questions and inquiries to Michael Hines or Ted Carnevale
Digital preprint of "The NEURON Simulation Environment" by M.L. Hines and N.T. Carnevale,
Neural Computation, Volume 9, Number 6 (August 15, 1997), pp. 1179-1209.
Copyright © 1997 by the Massachusetts Institute of Technology, all rights reserved. |